Similarities/differences between the porcupines of different Hemispheres: Erethizon vs Hystrix, part 1
@kevinatbrakputs @karoopixie @alexanderr @maxallen @caraleigh @danavan @jwidness @ken_j_allison @edporopat @arborsphere @whutchins @swampy @jnstuart @e-aus-kanada @nsharp @alwoodhouse @saber_animal @jakob @ariel-shamir @gwark @alanmuchlinski @bartwursten @markusgmeiner @christiaan_viljoen @markuslilje @opisska @ockertvs @lovecamp @asio-otus @lynnharper @birderryan @astanyoung @fatroosterfarm @miscelaineous @pfau_tarleton @donaldmcleod @pinawapt @jerry_deboer @douglasriverside @nvpyromelana @colincroft @jasonheadley @alexlamoreaux @aterrigeol @zonotrick @cindychrisler @tmurray74 @mlodinow @ungerlord @michael_oldham @desmond_macneal @camerondeckert @scottranger @justinhawthorne @jujurenoult @er1kksen @davidrscott @katzyna @ksled @dan_macneal @md-in-ns15 @mnerrie @dlcoleman @arcticory @nn4ever @codystricker @baywren @bert_raccoon @mikehannisian @meganhanson @amzapp @janeyd @mc1991 @josh_vandermeulen @zaccota @creedo @howlin_jay @erwinsieben @nyoni-pete @dewald2 @marco_vicariotto @emilianomori @ryruther @benjamynweil @ivanovdg19 @vmoser @natemartineau @lucaboscain @felix_riegel @mhughes26
(Many thanks to Kim Cabrera @beartracker for information useful in this series of Posts.)
Among the various mammals that defend themselves by means of spines, 'porcupines'
- are the largest-bodied forms (https://www.adn.com/alaska-news/science/2022/01/08/how-do-stagnant-bark-nibbling-porcupines-survive-frigid-alaska-winters/ and https://www.thedodo.com/bottle-fed-porcupine-returns-to-her-rescuers-1896274948.html),
have the most extreme forms of spines (https://www.naturalhistorymag.com/htmlsite/master.html?https://www.naturalhistorymag.com/htmlsite/0306/0306_feature.html and http://hotcore.info/babki/Porcupine-Quills-In-Dogs.html and
https://www.youtube.com/watch?v=ZphlCdI2yqA and https://www.jstor.org/stable/1383565)
and https://www.researchgate.net/publication/229674283_Mechanical_design_of_hedgehog_spines_and_porcupine_quills and https://www.researchgate.net/publication/341999648_Microstructure_of_Quills_in_Sunda_Porcupine_Hystrix_javanica_F_Cuvier_1823_Mikrostruktur_Duri_Landak_Jawa_Hystrix_javanica#:~:text=Nutritional%20content%20of%20the%20quills%20were%20water%20(89.93%25)%2C%20crude,sulphur%20(2%2C01%25),- possess spines even as infants (https://people.com/pets/baby-porcupine-born-in-front-of-guests-at-london-zoo/ and https://www.inaturalist.org/observations/102620880 and https://www.shutterstock.com/nb/image-photo/indian-crested-porcupine-baby-on-white-1038376888 and https://www.shutterstock.com/nb/image-photo/crested-porcupine-hystrix-cristata-female-young-1645995043),
- have warning colouration on both spines and other parts of the pelage (https://www.inaturalist.org/observations/46932604 and https://www.inaturalist.org/observations/89417614 and https://www.inaturalist.org/observations/80829627 and https://www.inaturalist.org/observations/80453530 and https://www.youtube.com/watch?v=woYjHBgm7oI), and
- have behaviours warning potential predators of the spiny defence, and retaliating to approach or contact with the well-developed guard hairs (for Erethizon, see https://www.alamy.com/porcupine-walking-through-a-the-summer-meadow-in-canada-image370342694.html and https://alaskafurid.wordpress.com/2009/11/02/porcupine/ and https://www.youtube.com/watch?v=-lgUmbhQ9vo and https://www.youtube.com/watch?v=cZ3UFsOlprI and https://www.youtube.com/watch?v=Z5iBzykfJko and https://www.youtube.com/watch?v=d28I9s6lpOY; for Hystrix see https://www.alamy.com/stock-photo-cutout-image-of-2-porcupine-quills-against-a-white-background-31305577.html).
However, the term 'porcupine' conflates two different lineages of rodents (https://animals.sandiegozoo.org/animals/porcupine). While these are similar enough to be confused in the public mind, they also differ significantly.
TAXONOMY
Any comparison is complicated by the fact that
there is only one species in Erethizon (https://academic.oup.com/mspecies/article/doi/10.2307/3504036/2600610?login=false and https://www3.uwsp.edu/biology/VertebrateCollection/Pages/Vertebrates/Mammals%20of%20Wisconsin/Erethizon%20dorsatum/Erethizon%20dorsatum.aspx#:~:text=Quills%20are%20modified%20guard%20hairs,pointing%20barbs%20(Roze%201989).
and https://www.chesapeakebay.net/discover/field-guide/entry/north_american_porcupine and http://www.adfg.alaska.gov/index.cfm?adfg=northamericanporcupine.video and https://www.turtlebay.org/north-american-porcupine and https://nhpbs.org/natureworks/porcupine.htm and https://animalsofpnw.com/2021/08/05/porcupines-of-the-pacific-northwest/), vs- five relevant species in Hystrix (excluding subgenus Thecurus, which is restricted to several islands in southeast Asia).
However, I treat collectively the five species of Hystrix, namely
- H. indica (mainland Asia, https://larastock.com/deposit-photo-221778628/?url=search/hystricomorph/ and https://www.istockphoto.com/photo/indian-crested-porcupine-on-grass-gm513787305-47371684 and https://rajajinationalpark.org/indian-crested-porcupine/ and https://www.projectnoah.org/spottings/1705106005/fullscreen and https://www.youtube.com/watch?v=Mk2RxZDb360 and https://www.tierpark-berlin.de/en/animals/indian-crested-porcupine and https://stock.adobe.com/search?k=%22indian+porcupine%22&asset_id=199150230),
- H. cristata (northern half of Africa plus Italy, https://escholarship.org/content/qt2tk1m81s/qt2tk1m81s_noSplash_3d0b3d6a295711913883e0eb5d1fe818.pdf?t=r1k9ob and https://sciencepress.mnhn.fr/sites/default/files/articles/pdf/az2010n2a2.pdf and https://www.mindenpictures.com/stock-photo-crested-porcupine-hystrix-cristata-foraging-on-ground-central-kenya-naturephotography-image90199652.html and https://www.shutterstock.com/nb/image-photo/porcupine-wildlife-africa-walking-around-thousand-142715425),
- H. africaeaustralis (southern half of Africa, https://www.sanbi.org/animal-of-the-week/cape-porcupine/ and https://www.inaturalist.org/observations/109172672 and https://focusedcollection.com/238091908/stock-photo-closeup-view-portrait-porcupine-running.html and https://www.alamy.com/stock-photo-cape-porcupine-or-south-african-porcupine-hystrix-africaeaustralis-24156097.html),
- H. brachyura (southeast Asia, https://upload.wikimedia.org/wikipedia/commons/5/54/Hystrix_brachyura%2C_Malayan_porcupine.jpg and https://www.shutterstock.com/nb/image-photo/profile-view-himalayan-porcupine-malayan-crested-2090406331 and https://www.shutterstock.com/nb/image-photo/malayan-porcupine-himalayan-large-porcupinehystrix-brachyura-525835324 and https://www.inaturalist.org/guide_taxa/999419 and https://www.ecologyasia.com/verts/mammals/malayan-porcupine.htm and https://naturerules1.fandom.com/wiki/Malayan_Porcupine?file=Malayan_Porcupine.jpg), and
- H. javanica (Indonesia, https://agris.fao.org/agris-search/search.do?recordID=DJ20210670575 and file:///C:/Users/Antoni%20Milewski/Downloads/3630-Article%20Text-6252-1-10-20190301.pdf and https://observation.org/photos/1414727/ and https://www.alamy.com/malayan-porcupine-in-cave-at-pangandaran-java-indonesia-image221010859.html and https://indonesiawindow.com/en/dozens-of-javanese-porcupines-return-to-natural-habitat/).
BODY SIZE and FORM
Erethizon and Hystrix overlap broadly in body mass, in the range 6-13 kg. However, the former is smaller-bodied than the latter.
Erethizon weighs as little as 3.5 kg when adult and lean, whereas Hystrix weighs as much as 25 kg. The heaviest, fattest specimens recorded for the two genera are 18 kg vs 27 kg.
In Erethizon, males outsize females; in Hystrix, the sexes have similar body size (https://www.tandfonline.com/doi/abs/10.1080/11250008709355592).
In https://www.youtube.com/watch?v=w4NfE9mqU64, the individual of Panthera pardus is an adult male, probably weighing more than 55 kg. By comparison, this individual of Hystrix africaeaustralis - as likely female as male - may weigh as much as 25 kg.
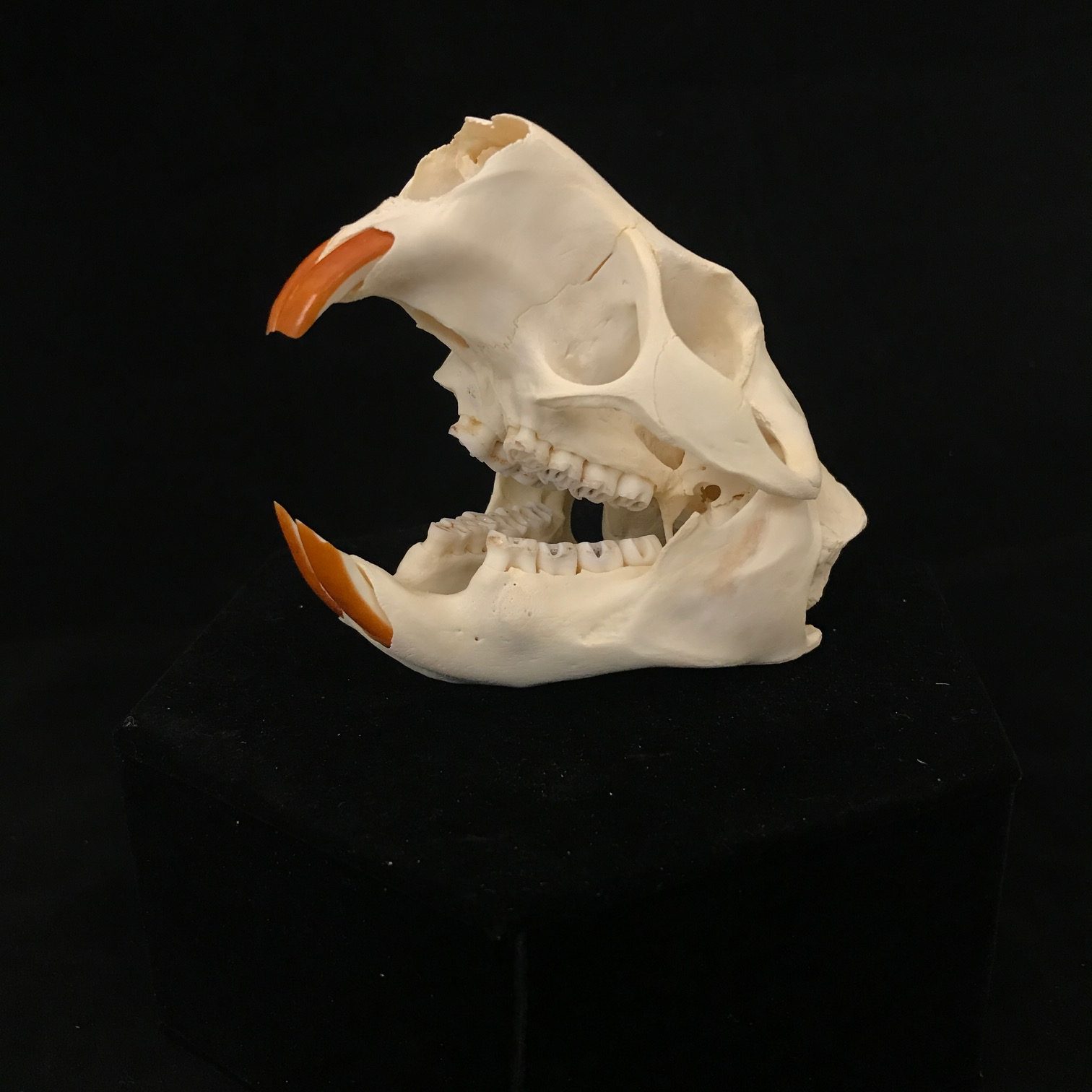
Both genera have small eyes (https://stock.adobe.com/sk/search/images?k=porcupine+face&asset_id=480402738 and https://stock.adobe.com/sk/search/images?k=porcupine+face&asset_id=175434829 and https://www.istockphoto.com/photo/crested-porcupine-gm90848177-2184836) and the same dental formula. However, the skulls differ surprisingly. That of Erethizon (https://animaldiversity.org/collections/contributors/skulls/erethizon/e._dorsatum/87345.lateral/) is unremarkable among rodents. By contrast, that of Hystrix (https://www.inaturalist.org/observations/36773744 and https://www.inaturalist.org/observations/17389159 and https://animaldiversity.org/collections/contributors/phil_myers/ADW_mammals/specimens/Rodentia/Hystricidae/Hystrix_indica/lateral6337/) has extremely inflated nasal and frontal bones.
The cranium also seem to differ in relative size. If Erethizon is brainier than Hystrix, this would be consistent with its play behaviour in adulthood (https://psycnet.apa.org/record/1944-03113-001) and its performance in memorising mazes (https://psycnet.apa.org/record/2005-14412-004).
The tail is proportionately larger in Erethizon (https://www.sciencephoto.com/media/1009962/view/north-american-porcupine-skeleton and https://fineartamerica.com/featured/1-north-american-porcupine-skeleton-millard-h-sharp.html and https://www.istockphoto.com/photo/north-american-porcupine-on-a-branch-gm519080646-90356241) than in Hystrix (https://paolov.files.wordpress.com/2015/03/20140227_143245.jpg and https://www.alamy.com/porcupine-skeleton-in-profile-view-after-antique-engraving-from-the-19th-century-image434355969.html).
POSTURES and LOCOMOTION
Both genera use the same gait in walking.
Erethizon:
https://www.inaturalist.org/observations/26949994 and https://www.shutterstock.com/nb/video/clip-2870482-north-american-porcupine-erethizon-dorsatum-walking-along and https://www.youtube.com/watch?v=cnos437-t18 and https://www.inaturalist.org/observations/95054372 and https://www.pond5.com/stock-footage/item/12263333-north-american-porcupine-walking-along-road-leaf-stuck-its-f.
Hystrix:
https://www.istockphoto.com/photo/porcupine-walking-gm177703348-24110328 and https://www.youtube.com/watch?v=5nbaSyg3Z1M.
Both genera prefer to commute along pathways, and Erethizon depends on creating its own pathways when commuting across deep snow.
The tracks of Erethizon (https://www.bear-tracker.com/porcpine.html) and Hystrix (https://twitter.com/bgsurvival/status/731435193278566400) are similar. In both genera, the feet are usually pointed somewhat inward in the walking gait.
However, the forefeet of the two genera differ. In Erethizon (https://natureidentification.com/how-to-identify-porcupine-erethizon-dorsatum-tracks/), the claws are long, the bare surface is rough, and the pollex is present, albeit only as a clawless pad. These features differ from Hystrix (https://www.natureinstock.com/search/preview/cape-porcupine-hystrix-africaeaustralis-adult-close-up-of-front-foot/0_11296644.html and https://www.inaturalist.org/observations/19891886).
Both Erethizon and Hystrix are able to swim (https://www.researchgate.net/publication/330451642_Behaviour_of_a_porcupine_Erethizon_dorsatum_swimming_across_a_small_boreal_stream and https://www.youtube.com/watch?v=wt1PMbVnB44). Neither genus seems capable of jumping.
However, postures and locomotion differ between the two genera in important ways.
Erethizon differs categorically from Hystrix in adopting the following postures/gaits, all of which use the muscular tail as a prop:
- sitting, which includes a hunched posture typical of the initial reaction to the approach of a potential predator (https://www.inaturalist.org/observations/14860384 and https://www.alamy.com/porcupine-standing-in-a-summer-meadow-in-canada-image370342894.html and https://www.peninsulaclarion.com/sports/the-porcupine-an-underappreciated-creature/ and https://www.dreamstime.com/porcupine-posing-his-hind-legs-spiky-porcupine-posing-his-hind-legs-giving-friendly-porcupine-smile-image128446819),
- standing and even walking bipedally (https://www.istockphoto.com/photo/north-american-porcupine-standing-on-hind-legs-erethizon-dorsatum-also-known-as-gm823905994-133351179 and https://www.vicnews.com/home2/photo-porcupine-taking-a-stroll-in-the-yukon-turns-heads/), for example during courtship, and
- climbing vertically (both upwards and downwards) by means of a gait in which the forelegs are extended in unison, with the hindlimbs moving subsequently in unison (https://www.youtube.com/watch?v=fXr7S3MbAaE and https://www.youtube.com/watch?v=w7R-GAUFqTg and https://www.inaturalist.org/observations/109799198).
Erethizon is capable of hanging upside down from branches (https://natlands.org/porcupines/ and https://www.istockphoto.com/photo/north-american-porcupine-gm1285939555-382593958), but cannot locomote while doing so.
By contrast, Hystrix climbs so poorly that adults can reputedly be kept captive by walls only one metre high.
Neither Erethizon nor Hystrix has spines on the ventral surfaces. However, in Erethizon the bristly hairs here - particularly on the ventral surface of the tail - enhance friction during climbing, compensating partly for the poor development of the pollex (https://www.inaturalist.org/observations/112849002 and https://www.shutterstock.com/nb/image-photo/north-american-porcupine-erethizon-dorsatum-on-310015790 and https://www.adfg.alaska.gov/index.cfm?adfg=northamericanporcupine.main).
Erethizon seems hardly able to run (https://www.youtube.com/watch?v=4v-XGIi0O4s and https://www.shutterstock.com/nb/image-photo/porcupine-running-thru-winter-snow-colville-108288905 and https://www.alamy.com/adult-north-american-porcupine-erethizon-dorsatum-running-in-glacier-bay-national-park-alaska-united-states-of-america-north-america-image417695139.html).
Hystrix is slow-moving relative to ungulates, but is versatile in its terrestrial gaits, as follows:
- it is capable of fleeing by trotting and cantering/galloping (https://www.youtube.com/watch?v=jl8nAJCKRUY and https://www.youtube.com/watch?v=tZnGBWB2Yew and https://www.shutterstock.com/nb/image-photo/crested-porcupine-running-across-meadow-1806708133 and https://www.youtube.com/watch?v=Blhyrwgq3j0 and https://www.youtube.com/watch?v=QVhTR9PGvSU), and
- its limb movements are rapid during the intermittent walking used in standoffs with potential predators; this defensive locomotion is forwards, backwards, and sideways according to the manoeuvres of the predator, so as to keep the longest spines and guard hairs pointed at the danger while repelling any close approach.
to be continued...


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}